As you might know, proteins are one of the major “building blocks” of cells; there’s up to 10.000 different types of proteins, all manufactured inside each cell. Abiogenesis theorists obviously supports the view that these molecules have arisen “by chance”, in a prebiotic world, billion years ago, however, to date, they have absolutely no clue about it, as we can read from this article:
“Proteins are the most complex chemicals synthesized in nature and must fold into complicated three-dimensional structures to become active. This poses a particular challenge in explaining their evolution from non-living matter. So far, efforts to understand protein evolution have focused on domains, independently folding units from which modern proteins are formed. Domains however are themselves too complex to have evolved de novo in an abiotic environment. We think that domains arose from the fusion of shorter, non-folding peptides, which evolved as cofactors supporting a primitive, RNA-based life form (the ‘RNA world’).” 1
So, why is it so complicated to explain its origin? Despite the often repeated innuendo that life and all of its components has “assuredly” originated through natural means, the clear failure of scientists to solve this puzzle can be easily explained by some truths about proteins, its synthesis, structure and so on. After that, no one can reasonably take its abiogenetic origin as logically granted. These truths also explain without shadow of doubt the intriguing fact that absolutely no single protein (even the lesser one, composed of only 8 amino acids) has ever been observed to appear anywhere in the world, outside the cells and high-tech labs, of course!
What’s a protein?
“Proteins are large biological molecules consisting of one or more chains of amino acids. Proteins perform a vast array of functions within living organisms, including catalyzing metabolic reactions, replicating DNA, responding to stimuli, and transporting molecules from one location to another. Proteins differ from one another primarily in their sequence of amino acids, which is dictated by the nucleotide sequence of their genes, and which usually results in folding of the protein into a specific three-dimensional structure that determines its activity.
A polypeptide is a single linear polymer chain of amino acids bonded together by peptide bonds between the carboxyl and amino groups of adjacent amino acid residues. The sequence of amino acids in a protein is defined by the sequence of a gene, which is encoded in the genetic code. In general, the genetic code specifies 20 standard amino acids;” 2
Talking about amino acids, we’d like to recall another crucial problem for abiogenesis: The absence of self-occurring homochiral mixtures. As it has been told in a previous article, the laws of thermodynamics obliges the occurrence of racemic mixtures, ever:
“The left and right handed forms have identical free energy (G), so the free energy difference (ΔG) is zero. The equilibrium constant for any reaction (K) is the equilibrium ratio of the concentration of products to reactants. The relationship between these quantities at any Kelvin temperature (T) is given by the standard equation:
K = exp (–ΔG/RT)
where R is the universal gas constant (= Avogadro’s number x Boltzmann’s constant k) = 8.314 J/K.mol.
For the reaction of changing left-handed to right-handed amino acids (L → R), or the reverse (R → L), ΔG = 0, so K = 1. That is, the reaction reaches equilibrium when the concentrations of R and L are equal; that is, a racemate is produced.”
Therefore, any abiogenetic theorist has this astounding problem to deal with from the very beginning; without homochiral monomers, we can have zero possibility of a ‘magic’ protein self-assembling…
Protein synthesis
It’s quite uncanny that intelligent people with advanced knowledge on the subject might attempt to conceive hypothesis of such molecules originating spontaneously, in the wild and morbid inorganic environment, because for cells to build proteins, an intricate, complex and laborious process must take place!
First, genetic information is needed:
“Proteins are assembled from amino acids using information encoded in genes. Each protein has its own unique amino acid sequence that is specified by the nucleotide sequence of the gene encoding this protein. The genetic code is a set of three-nucleotide sets called codons and each three-nucleotide combination designates an amino acid (for example AUG (adenine–uracil–guanine) is the code for methionine).”
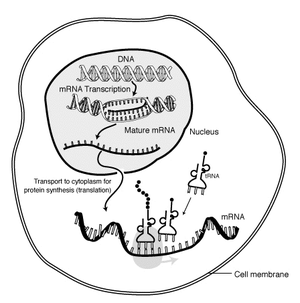
Many proteins use more that one of the 64 possible codons to be built. Moreover, that specific genetic code must be first translated, transcribed:
“Genes encoded in DNA are first transcribed into pre-messenger RNA (mRNA) by proteins such as RNA polymerase. Most organisms then process the pre-mRNA (also known as a primary transcript) using various forms of Post-transcriptional modification to form the mature mRNA, which is then used as a template for protein synthesis by the ribosome.”
Oh great, a bit complicated, isn’t it? Please, read the Wikipedia article referring to the messenger RNA, for further comprehension of what it is, its manufacturing, composition, etc; all of which adds up more complexity for the protein origin’s explanation.
The process of synthesizing a protein from an mRNA template is known as translation. The mRNA is loaded onto the ribosome and is read three nucleotides at a time by matching each codon to its base pairinganticodon located on a transfer RNA molecule, which carries the amino acid corresponding to the codon it recognizes. The enzyme aminoacyl tRNA synthetase “charges” the tRNA molecules with the correct amino acids. The growing polypeptide is often termed the nascent chain. Proteins are always biosynthesized from N-terminus to C-terminus.[6]
The size of a synthesized protein can be measured by the number of amino acids it contains and by its total molecular mass, which is normally reported in units of daltons (synonymous with atomic mass units), or the derivative unit kilodalton (kDa). Yeast proteins are on average 466 amino acids long and 53 kDa in mass.[5]The largest known proteins are the titins, a component of the musclesarcomere, with a molecular mass of almost 3,000 kDa and a total length of almost 27,000 amino acids.[8]
Phew! How complicated! You may ask now: are we finally done? And I reply you: Huh, nope! Now that the ribosome, together with the rRNA and more than 50 other proteins, has finally finished the process, a protein is formed. However, it is always found in a random coil shape. So what? This shape is mostly useless for its usage on organism, as we can read:
Each protein exists as an unfolded polypeptide or random coil when translated from a sequence of mRNA to a linear chain of amino acids. This polypeptide lacks any stable (long-lasting) three-dimensional structure (the left hand side of the neighbouring figure). 3
In that randomly coiled shape, the protein is highly unstable, breakable, useless for cell building, so, for proper biological use and better stability, the protein folding process must take place. This 3D-shape is known as the native state.
The correct three-dimensional structure is essential to function, although some parts of functional proteins may remain unfolded.[4] Failure to fold into native structure generally produces inactive proteins, but in some instances misfolded proteins have modified or toxic functionality. Several neurodegenerativeand other diseases are believed to result from the accumulation of amyloid fibrils formed by misfolded proteins.[5] Many allergies are caused by incorrect folding of some proteins, for the immune system does not produce antibodies for certain protein structures.[6]
Another importance of the protein folding is:
Minimizing the number of hydrophobic side-chains exposed to water is an important driving force behind the folding process.[9] Formation of intramolecular hydrogen bonds provides another important contribution to protein stability.[10]
The process of folding often begins co-translationally, so that the N-terminus of the protein begins to fold while the C-terminal portion of the protein is still beingsynthesized by the ribosome. Specialized proteins called chaperones assist in the folding of other proteins.
Although most globular proteins are able to assume their native state unassisted, chaperone-assisted folding is often necessary in the crowded intracellular environment to prevent aggregation; chaperones are also used to prevent misfolding and aggregation that may occur as a consequence of exposure to heat or other changes in the cellular environment.
There are two models of protein folding that are currently being confirmed: The first: The diffusion collision model, in which a nucleus is formed, then the secondary structure is formed, and finally these secondary structures are collided together and pack tightly together. The second: The nucleation-condensation model, in which the secondary and tertiary structures of the protein are made at the same time. Recent studies have shown that some proteins show characteristics of both of these folding models.
The essential fact of folding, however, remains that the amino acid sequence of each protein contains the information that specifies both the native structure and the pathway to attain that state. Folding is a spontaneous process independent of energy inputs from nucleoside triphosphates. The passage of the folded state is mainly guided by hydrophobic interactions, formation of intramolecular hydrogen bonds, and van der Waals forces, and it is opposed by conformational entropy.
Only after the folding process, we have an useful, stable protein, with a properly designed shape with its up to four layers, so that the molecule can perform its biological function.
But, remember, many conditions and external factors can destroy proteins, such as hydrolysis (it’s a slow, but ceaseless process, because proteins are metastable, hydrophobic) and others:
Under some conditions proteins will not fold into their biochemically functional forms. Temperatures above or below the range that cells tend to live in will cause thermally unstableproteins to unfold or “denature” (this is why boiling makes an egg white turn opaque). High concentrations of solutes, extremes of pH, mechanical forces, and the presence of chemical denaturants can do the same.
A fully denatured protein lacks both tertiary and secondary structurel. Under certain conditions some proteins can refold; however, in many cases, denaturation is irreversible.[15] Cells sometimes protect their proteins against the denaturing influence of heat with enzymes known as chaperones or heat shock proteins, which assist other proteins both in folding and in remaining folded. Some proteins never fold in cells at all except with the assistance of chaperone molecules, which either isolate individual proteins so that their folding is not interrupted by interactions with other proteins or help to unfold misfolded proteins, giving them a second chance to refold properly. This function is crucial to prevent the risk of precipitation into insoluble amorphous aggregates.
For a further an in-depth study about different factors capable of disrupting proteins, read the following articles:
To conclude our observation, it’s impossible not to be sceptic of any theoretic proposition that claims self-caused origin of proteins, because it turns out that science unveiled tons of facts that easily prevent any possibility of such proposed scenario:
-Absence of homochiral monomers forming in the environment;
-Necessity of genetic specific information;
-Need for an highly controlled ambient, with proper Ph level, temperature, absence of mechanical forces that may easily damage, disrupt the protein, toxins, etc;
-Need for specific methods to protect the protein against hydrolysis, oxidation;
-Necessity of having 50 other types of protein already manufactured to help on the protein synthesis;
The question raises: how in the world could such a specific set of conditions be found in a prebiotic Earth? Such condition can only be barely found in a first-class laboratory, driven by qualified and experienced scientists!
You might as well enjoy watching this short video talking about protein synthesis:
Glycolysis (from glycose, an older term for glucose + -lysis degradation) is the metabolic pathway that converts glucose C6H12O6, into pyruvate, CH3COCOO− + H+. The free energy released in this process is used to form the high-energy compounds ATP (adenosine triphosphate) and NADH (reduced nicotinamide adenine dinucleotide).Yeast cells obtain energy under anaerobic conditions using a very similar process called alcoholic fermentation, also referred to as ethanol fermentation, is a biological process in which sugars such as glucose, fructose, and sucrose are converted into cellular energy and thereby produce ethanol and carbon dioxide as metabolic waste products.
Glycolysis requires 11 enzymes which degrade glucose to lactic acid (Fig. 2). Alcoholic fermentation follows the same enzymatic pathway for the first 10 steps. The last enzyme of glycolysis, lactate dehydrogenase, is replaced by two enzymes in alcoholic fermentation. These two enzymes, pyruvate decarboxylase and alcoholic dehydrogenase, convert pyruvic acid into carbon dioxide and ethanol in alcoholic fermentation.
The most commonly accepted evolutionary scenario states that organisms first arose in an atmosphere lacking oxygen.1,2 Anaerobic fermentation is supposed to have evolved first and is considered the most ancient pathway for obtaining energy. However, there are several scientific odds against that.
First of all, it takes ATP energy to start the process that will only later generate a net gain in ATP. Two ATPs are put into the glycolytic pathway for priming the reactions, the expenditure of energy by conversion of ATP to ADP being required in the first and third steps of the pathway (Fig. 2). A total of four ATPs are obtained only later in the sequence, making a net gain of two ATPs for each molecule of glucose degraded. The net gain of two ATPs is not realized until the tenth enzyme in the series catalyzes phosphoenolpyruvate to ATP and pyruvic acid (pyruvate). This means that neither glycolysis nor alcoholic fermentation realizes any gain in energy (ATP) until the tenth enzymatic breakdown.
Enzymes are proteins consisting of amino acids united in polypeptide chains. Their complexity may be illustrated by the enzyme glyceraldehyde 3-phosphate dehydrogenase, which is the enzyme that catalyzes the oxidation of phosphoglyceraldehyde in glycolysis and alcoholic fermentation. Glyceraldehyde phosphate dehydrogenase consists of four identical chains, each having 330 amino acid residues. The possible number of different combinations of these amino acid chains is infinite.
Glyceraldehyde-3-phosphate dehydrogenase
To illustrate, let us consider a simple protein containing only 100 aim acids. There are 20 different kinds of L-amino acids in proteins, and each can be used repeatedly in chains of 100. Therefore, they could be arranged in 20^100 or 10^130 different ways. Even if a hundred million billion of these (10^17) combinations could function for a given purpose, there is only one chance in 10^113 of getting one of these required amino acid sequences in a small protein consisting of 100 amino acids. By comparison, Sir Arthur Eddington has estimated there are no more than 10^80 (or 3,145 x 10^79) particles in the universe! Consider the 10 enzymes of the glycolytic pathway. If each of these were a small protein having 100 amino acid residues with some flexibility and a probability of 1 in 10^113 or 10^-113, the probability for arranging the amino acids for the 10 enzymes would be: P = 10^-1,130 or 1 in 10^1,130, and this result is only the odds against producing the 10 glycoytic enzymes by chance. It is estimated that the human body contains 25,000 enzymes. If each of these were only a small enzyme consisting of 100 amino acids with a probability of 1 in 10^-113, the probability of getting all 25,000 would be (10^-113)^25,000, which is 1 chance in 10^2,825,000…
There are still other problems with that theory. There are numerous complex regulatory mechanisms which control these chemical pathways. For example, phosphofructokinase is a regulatory enzyme which limits the rate of glycolysis. Glycogen phosphorylase is also a regulatory enzyme; it converts glycogen to glucose-1-phosphate and thus makes glycogen available for glycolytic breakdown. In complex organisms there are several hormones such as somatotropin, insulin, glucagon, glucocorticoids, adrenaline thyroxin and a host of others which control utilization of glucose.
In addition, complex cofactors are absolutely essential for glycolysis. One of the two key ATP energy harvesting steps in glycolysis requires a dehydrogenase enzyme acting in concert with the “hydrogen shuttle” redox reactant, nicotinamide adenine dinucleotide (NAD+). To keep the reaction sequence going, the reduced cofactor (NADH + H +) must be continuously regenerated by steps later in the sequence (Fig. 2), which requires one enzyme in glycolysis (lactic dehydrogenase) and another (alcohol dehydrogenase) in alcoholic fermentation.
Further, at one point, an intermediate in the glycolytic pathway is “stuck” with a phosphate group (needed to make ATP) in the low energy third carbon position. A remarkable enzyme, a “mutase” (Step 8), shifts the phosphate group to the second carbon position—but only in the presence of pre-existent primer amounts of an extraordinary molecule, 2,3-diphosphoglyceric acid. Actually, the shift of the phosphate from the third to the second position using the “mutase” and these “primer” molecules accomplishes nothing notable directly, but it “sets up” the ATP energy-harvesting reaction which occurs two steps later!
by Jean Sloat Morton, Ph.D.
References
1 A.I. Oparin, Origin of Life, New York: Dover Pub., lnc., 1965, pp. 225-26.
2 (Jark and Synge (eds.), The Origin of Life on the Earth, New York: Pergamon Press, 1959, p. 52.
3 Ernil Borel, Probabilities and Life, New York: Dover Pub., Inc., 1962, p. 28.
Cite this article: Morton, J. S. 1980. Glycolysis and Alcoholic Fermentation. Acts & Facts. 9 (12).
Although water is portrayed by many as the solution to, or star player in how life came to be, the fact is that water spontaneously breaks down complex molecules that living organisms need to exist: such as DNA,* RNA, proteins and their components.** For example, an article on Molecular Cloning says that “Proteins are usually soluble in water solutions because they have hydrophilic amino acids on their surfaces.”1
Amino acids have been called the building blocks of life, and when two or more are joined together they are called a peptide and the bond that holds them together is called a peptide bond. When ten or more are linked together they may be called a polypeptide, and if they are ordered and folded correctly, they become a protein. And in a wikipedia article on peptide bonds we are told that “a peptide bond can be broken by … hydrolysis” *** (just by) … “adding … water”… (and that the) “… bonds in proteins are metastable, meaning that in the presence of water they will break spontaneously.”2
A simple video explain dehydratation and hydrolysis:
Another article on this topic 3 says that hydrolysis is:
“A chemical reaction in which water is used to break the bonds of certain substances.In biotechnology and living organisms, these substances are often polymers …such as that … (exist) between two amino acidsin aprotein … “
Dr. A. E. Wilder-Smith, (Ph.D. organic chemistry) also brought this out in a book he wrote on life’s complexity and origin.4
“Amino acids and other building blocks present in the macromolecules of living matter aggregate to form larger units … by … (a reaction) called condensation.**** The combinations usually involve the elimination of one molecule of water between two combining molecules. It is the removal of this molecule of water which presents the major difficulty … For, the removal of this water molecule from between two combining molecules requires energy which must … be supplied in some fashion.
“A further difficulty arises in this question of the elimination of water. For, in the prebiotic world, it is assumed that the condensation reaction took place in the presence of a large … (supply) of water which would tend, according to the law of mass action, to hinder the condensation process and … (promote)decomposition (or breakdown of peptides and polypeptides). … The more water, the less condensation.”
“If the reaction is to proceed in the direction of the dipeptide, (or two amino acids that are joined together) … the water molecule … (that results) must be removed from the reaction system since the reaction is reversible. If it is not removed … (it will) hydrolyze (or separate) the dipeptide back again to the constituent amino acids …”
This means the “primordial soup,”or “warm little pond” where Darwin speculated that life began could not have been simply water, since it would “hydrolyze” or break down complex molecules back into their basic original amino acid as soon as they formed. Dr. Charles McCombs explains the problem as follows in an article he wrote on the subject of why life by chance is virtually, if not utterly and completely impossible.
“Every time one component reacts with a second component forming the polymer, the chemical reaction also forms water as a byproduct … There is a rule of chemical reactions … called the Law of Mass Action that says all reactions proceed in a direction from highest to lowest concentration. This means that any reaction that produces water cannot be performed in the presence of water. This Law of Mass Action provides a total hindrance to protein, DNA/RNA, and polysaccharide formation because even if the condensation took place, the water from a supposed primordial soup would immediately hydrolyze them. Thus, if they are formed according to evolutionary theory, the water would have to be removed … which is impossible in a “watery” soup.5
But because the “watery soup” in living cells is surrounded by a membrane, the “water” inside the cell “behaves very differently” 6 than ordinary water. In fact, the “water” in a cell is not water but a blend of water, amino acids, proteins, and many other chemicals called cytosol. This mixture is the result of the DNA’s ability to regulate what goes in and out of the cell — via numerous channels that control and regulate what is allowed to pass through the cell membrane, and thus to create and maintain a favorable environment and PH for DNA, RNA and protein synthesis, and life itself to exist.
If the concentration of amino acids is high enough, some of them will link up with others to form dipeptides and tripeptides. An article on this subject states that:
“is important to recognize that by whatever reactions polymerization (or the joining of amino acids)occurred, they had to be reactions that would occur in an essentially aqueous environment. This presents difficulties because condensation of amino acids to form peptides, or of nucleotides to form RNA or DNA, is not thermodynamically favorable in aqueous solution.”{6}
The explanation for this is partly that the concentration of amino acids decreases as amino acids form pairs (called dipeptides) in a solution. This decreased concentration causes the velocity of the peptide synthesis reaction to slow down, and some dipeptides begin breaking up, again becoming single amino acids. The solution reaches equilibrium when just as many dipeptides dissociate as associate. A very tiny fraction of the dipeptides add another amino acid to form a tripeptide. … Oligopeptides (Oligo=few) and polypeptides (poly=many) will form only very rarely. Tripeptides dissociate faster than dipeptides in the same solution. 7
In this regard, a tripeptide has only three amino acids, while the simplest protein ever found has at least eight: connected in a specific order.
Jeffrey P. Tomkins makes the following statement in a book on the design and complexity of the cell:
“… plasma membranes are … quite complex and … (function) as more than just a barrier … Some key functions of the membrane involve the import and export of chemical compounds through specialized transmembrane channels, sensory and signaling processes via specialized receptor proteins imbedded in the membrane, and osmotic (water) regulation … through special portals.” 8
“Within the … membrane is the internal cell matrix … called cytosol or cytoplasm, which is a semi-fluid substance. … Like the … membrane, the complexity of … cytoplasm seems to grow with every new discovery in cell biology.” 8
Tomkins also tells us that water must be regulated and controlled outside the cell as well in what is called the “extra cellular matrix.” 8
This means that the water of yesteryear, or the distant past, almost certainly performed just like the water of today, and that water, dirt and chemicals, could not have created life anymore than fuel, dirt, and metallic ore, — by themselves — could create a car, motorcycle, or an airplane: even in millions, billions, or trillions of years.
A typical human cell would undergo 2,000 to 10,000 spontaneous DNA hydrolysis damage events every day just because it is an aqueous environment.(The only reason that DNA functions as well as it does is that cells come equipped with an amazing array of cooperative DNA repair mechanisms. For example, polymerase replication during cell division might produce 6 million errors per cell, but then proofreading machinery can reduce this to 10,000 and then mis-match repair machinery could reduce this to 100. It appears to be impossible, however, to replicate the 6 billion nucleotides in a human cell in a completely error-free manner.
A way to remove water is with certain high-energy chemicals that absorb water, called condensing agents. If the reaction between condensing agent C and water is:
C + H2O → D (2)
and if ΔG2 of reaction (2) is negative and large enough, it can couple with reaction (1):
H2NCHRCOOH + H2NCHR′COOH + C → H2NCHRCONHCHR′COOH + D (3)
ΔG3 = ΔG1 + ΔG2. If ΔG3 is large and negative, the equilibrium constant for reaction 3, K3, will be large, and this could conceivably produce reasonable quantities of polymers.
Some researchers used the condensing agent dicyanamide (N=CNHC=N) to produce some peptides from glycine, even claiming, ‘dicyanamide mediated polypeptide synthesis may have been a key process by which polypeptides were produced in the primitive hydrosphere.’
However, the biggest problem is that condensing agents would readily react with any water available. Therefore it is a chemical impossibility for the primordial soup to accumulate large quantities of condensing agents, especially if there were millions of years for water to react with them. Yet the Sydney Fox experiment used a 30-fold excess of dicyanamide. And even with these unrealistic conditions, 95% of the glycine remained unreacted, and the highest polymer formed was a tetrapeptide.
* Although chemists can make DNA in their laboratories, they can only do so under highly controlled conditions that simulate cytosol. They achieve this by using a pre-existing DNA or gene (template), using the right amount of water, magnesium chloride, and salt buffers, and by using a pre-existing microscopic / molecular copy machine called DNA polymerase. Such would not be the case in nature, since genes are not known to form by themselves, nor even simple proteins that consist of only 8 amino acids: much less complex ones that consist of 900–1000 of them, such as DNA polymerase — along with a motor protein called helicase: that actually spins like a motor (at 1800 rpms) and that unwinds the DNA.
** When two amino acids come together they are called a peptide, and the reaction is called condensation or a condensation reaction **** or dehydration synthesis. A nucleic acid is a synonym for a nucleotide, and when two or more nucleotides join together they are called an oligonucleotide.
*** According to the American Heritage Dictionary of Science, hydrolysis is “a process of decomposition in which a compound is broken down and changed into other compounds by … (absorbing, or being diluted with) water. For example, in food digestion, the food absorbs water and is broken down by hydrolysis. The same dictionary says that to hydrolyze means “to decompose by hydrolysis …” and that organic molecules such as “Nucleic acids, proteins, and polysaccharides contain many bonds that hydrolyze … In this regard, the combining word hydro- simply means “of or having to do with water.”
**** Think of a Condensed can of Campbell’s Soup. The fact that it is “condensed” simply means that water has been removed.
Deoxyribonucleic acid (DNA) is a molecule that encodes the genetic instructions used in the development and functioning of all known living organisms and many viruses. Along with RNA and proteins, DNA is one of the three major macromolecules essential for all known forms of life. Most DNA molecules are double-stranded helices, consisting of two long biopolymers of simpler units called nucleotides—each nucleotide is composed of a nucleobase (guanine, adenine, thymine, and cytosine), recorded using the letters G, A, T, and C, as well as a backbone made of alternating sugars (deoxyribose) and phosphate groups (related to phosphoric acid), with the nucleobases (G, A, T, C) attached to the sugars. DNA is well-suited for biological information storage, since the DNA backbone is resistant to cleavage and the double-stranded structure provides the molecule with a built-in duplicate of the encoded information. The following video shows animations of processes which occur within each one of the 10 trillion cells in our body!
Discovery
In 1927, Nikolai Koltsov proposed that inherited traits would be inherited via a “giant hereditary molecule” made up of “two mirror strands that would replicate in a semi-conservative fashion using each strand as a template”. In 1928, Frederick Griffith discovered that traits of the “smooth” form of Pneumococcus could be transferred to the “rough” form of the same bacteria by mixing killed “smooth” bacteria with the live “rough” form. This system provided the first clear suggestion that DNA carries genetic information—the Avery–MacLeod–McCarty experiment—when Oswald Avery, along with coworkers Colin MacLeod and Maclyn McCarty, identified DNA as the transforming principle in 1943. DNA’s role in heredity was confirmed in 1952, when Alfred Hershey and Martha Chase in the Hershey–Chase experiment showed that DNA is the genetic material of the T2 phage.
In 1953, James Watson and Francis Crick suggested what is now accepted as the first correct double-helix model of DNA structure in the journal Nature. Their double-helix, molecular model of DNA was then based on a single X-ray diffraction image (labeled as “Photo 51”) taken by Rosalind Franklin and Raymond Gosling in May 1952, as well as the information that the DNA bases are paired — also obtained through private communications from Erwin Chargaff in the previous years. Chargaff’s rules played a very important role in establishing double-helix configurations for B-DNA as well as A-DNA.
Experimental evidence supporting the Watson and Crick model was published in a series of five articles in the same issue of Nature. Of these, Franklin and Gosling’s paper was the first publication of their own X-ray diffraction data and original analysis method that partially supported the Watson and Crick model; this issue also contained an article on DNA structure by Maurice Wilkins and two of his colleagues, whose analysis and in vivo B-DNA X-ray patterns also supported the presence in vivo of the double-helical DNA configurations as proposed by Crick and Watson for their double-helix molecular model of DNA in the previous two pages of Nature.
Repairing system
It’s interesting to notice that the DNA molecule is highly reactive, thus, very unstable… On a good day about one million bases in the DNA in a human cell are damaged. These lesions are caused by a combination of normal chemical activity within the cell and exposure to radiation and toxins coming from environmental sources including cigarette smoke, grilled foods and industrial wastes. So, the organisms have a handful of repairing mechanisms, as said in a recent Science Daily article:
“A number of environmental toxins and chemotherapy drugs are alkylation agents that can attack DNA.
When a DNA base becomes alkylated, it forms a lesion that distorts the shape of the molecule enough to prevent successful replication. If the lesion occurs within a gene, the gene may stop functioning. To make matters worse, there are dozens of different types of alkylated DNA bases, each of which has a different effect on replication.
One method to repair such damage that all organisms have evolved is called base excision repair. In BER, special enzymes known as DNA glycosylases travel down the DNA molecule scanning for these lesions. When they encounter one, they break the base pair bond and flip the deformed base out of the DNA double helix. The enzyme contains a specially shaped pocket that holds the deformed base in place while detaching it without damaging the backbone. This leaves a gap (called an “abasic site”) in the DNA that is repaired by another set of enzymes.
Human cells contain a single glycosylase, named AAG, that repairs alkylated bases. It is specialized to detect and delete “ethenoadenine” bases, which have been deformed by combining with highly reactive, oxidized lipids in the body. However, AAG also handles many other forms of akylation damage. Many bacteria, however, have several types of glycosylases that handle different types of damage.
“It’s hard to figure out how glycosylases recognize different types of alkylation damage from studying AAG since it recognizes so many,” says Eichman. “So we have been studying bacterial glycosylases to get additional insights into the detection and repair process.”
That is how they discovered the bacterial glycosylase AlkD with its unique detection and deletion scheme. All the known glycosylases work in basically the same fashion: They flip out the deformed base and hold it in a special pocket while they excise it. AlkD, by contrast, forces both the deformed base and the base it is paired with to flip to the outside of the double helix. This appears to work because the enzyme only operates on deformed bases that have picked up an excess positive charge, making these bases very unstable. If left alone, the deformed base will detach spontaneously. But AlkD speeds up the process by about 100 times. Eichman speculates that the enzyme might also remain at the location and attract additional repair enzymes to the site.
AlkD has a molecular structure that is considerably different from that of other known DNA-binding proteins or enzymes. However, its structure may be similar to that of another class of enzymes called DNA-dependent kinases. These are very large molecules that possess a small active site that plays a role in regulating the cells’ response to DNA damage. AlkD uses several rod-like helical structures called HEAT repeats to grab hold of DNA. Similar structures have been found in the portion of DNA-dependent kinases with no known function, raising the possibility that they play an additional, unrecognized role in DNA repair.”
It’s impossible to conceive that such unstable, complex molecule could have originated all alone, by chance and lasted any long in a pre-biotic environment. In a primordial Earth, with a free-oxygen atmosphere, it turns out that it would have no ozon layer, or a very thin one. Thus, the UV light from the Sun would freely bombards the Earth without filtering; the most damaging UV light types would face no barrier, and it’s a fact that UV light damages, degrades polymers!
Many natural and synthetic polymers are attacked by ultra-violet radiation and products made using these materials may crack or disintegrate (if they’re not UV-stable). The problem is known as UV degradation, and is a common problem in products exposed to sunlight. Continuous exposure is a more serious problem than intermittent exposure, since attack is dependent on the extent and degree of exposure.
Effect of UV exposure on polypropylene rope (the left one is damaged, the right one is a new rope)
Add to it the oxidation problem, hydrolysis, etc… Any natural origin of RNA/DNA is inconceivable!